Catalogue of Palaearctic species

Two brood cells of Hoplitis (Anthocopa) villosa (Schenck, 1853) in a cavity between stones; the brood cells are constructed from petals, which are glued together with varying amounts of mud. Foto F. Amiet.
The species accounts contain the (1) taxonomy, (2) the geographic distribution, (3) literature sources for species identification as well as a short summary of our current knowledge on (4) nesting behaviour and (5) flower preferences.
Taxonomy
The species-group names are arranged alphabetically by genus and subgenus, and chronologically within each valid name. The agreement of gender of the species-group name with the generic name as requested by the Article 31.2 of the Code of Zoological Nomenclature is enforced throughout the catalogue and hence the gender ending is changed if necessary. Griswold and Michener (1998) and Michener (2007) were used for the placement of the species-group taxa into the accepted genus-group taxa. We slightly deviate from this system by following European authors in their circumscription of Osmia (Allosmia) versus Osmia (Erythrosmia). In addition, we keep Hoplitis (Micreriades) as a distinct subgenus separated from Hoplitis (Alcidamea) and treat Nasutosmia as a subgenus of Osmia rather than of Hoplitis. Furthermore, Stenosmia and Hoplosmia are reduced to subgeneric rank in Hoplitis and Osmia, respectively, and Pseudoheriades is removed from the osmiine bees. These deviations are justified by the results of a molecular phylogenetic study of the Osmiini (Praz et al., 2008b). In addition, i) Ceraheriades, a former subgenus of Chelostoma, is treated here as junior synonym of Prochelostoma based on a recent phylogenetic analysis (Sedivy et al., 2008), ii) Exanthocopa, a former subgenus of Hoplitis, is synonymized with the subgenus Anthocopa (A. Müller, unpublished), iii) Ozbekosmia, a former subgenus of Osmia, is synonymized with the subgenus Tergosmia (A. Müller, unpublished), iv) the subgenera Annosmia, Bytinskia, Coloplitis and Hoplitis are merged into one large subgenus Hoplitis, v) the subgenera Alcidamea, Megalosmia, Monumetha and Prionohoplitis are merged into one large subgenus Alcidamea based on a recent molecular phylogenetic analysis (Sedivy et al., 2013a, c), and vi) the subgenera Monosmia and Orientosmia are synonymized with the subgenus Osmia based on a combined molecular and morphological phylogeny (Haider et al., 2013). All alternative published opinions concerning synonymy, rank as well as generic and subgeneric placement are disclosed citing at least one reference (not necessarily the earliest). Extralimital genera of the Osmiini restricted to areas outside the Palaearctic and therefore not treated here are Ashmeadiella Cockerell, Atoposmia Cockerell, Bekilia Benoist, Othinosmia Michener and Xeroheriades Griswold.
Symbols
†extinct taxon
# original publication of a name not seen by the authors of the catalogue
m/f one male/female specimen
mm/ff several male/female specimens
m(m)/f(f) male(s)/female(s) of unknown quantity
CH country acronym for type localities
CH? country acronym for uncertain localities
CH country acronym for recorded localities which have been shown to be demonstrably incorrect by subsequent authors
[…] note by the authors of this catalogue
[= …] currently accepted combination of a type species
Collections
AMNH American Museum of Natural History, New York United States
BMNH The Natural History Museum, London United Kingdom
DAAN Department of Agriculture, Ministry of Agriculture and Natural Resources, Nicosia Cyprus
DEI Deutsches Entomologisches Institut, Müncheberg Germany
DEZA Dipartimento di Entomologia e Zoologia Agraria dell’Università, Portici Italy
EMET Ataturk Universitesi, Faculty of Agriculture, Entomology Museum, Erzurum Turkey
ETHZ Eidgenössische Technische Hochschule, Entomologische Sammlung, Zürich Switzerland
FMNH Finnish Museum of Natural History, Helsinki Finland
GMUG Universität Göttingen, Geowissenschaftliches Zentrum, Göttingen Germany
HNHM Hungarian Natural History Museum, Budapest Hungary
IEGG Università di Bologna, Istituto di Entomologia, Bologna Italy
ISZP Polish Academy of Sciences, Institute of Systematic Zoology, Krakow Poland
IZCAS Chinese Academy of Sciences, Institute of Zoology, Beijing China
KUEC Kyushu University, Fukuoka Japan
LSUK Linnean Society, London United Kingdom
MHNG Muséum d’Histoire Naturelle, Genève Switzerland
MICN Museo Insular de Ciencias Naturales, Canary Islands, Tenerife Spain
MLUH Martin-Luther-Universität, Wissenschaftsbereich Zoologie, Halle Germany
MNHN Muséum Nationale d’Histoire Naturelle, Paris France
MSDB Museo di Storia Naturale, Torino Italy
MSNG Museo Civico di Storia Naturale, Genova Italy
MTD Museum für Tierkunde, Dresden Germany
MZLS Musée Zoologique, Lausanne Switzerland
MZLU Lund University, Lund Sweden
NHMB Naturhistorisches Museum, Basel Switzerland
NHRS Swedish Museum of Natural History, Stockholm Sweden
NMBE Naturhistorisches Museum, Bern Switzerland
NMPC National Museum (National History), Prague Czech Republic
NMW Naturhistorisches Museum, Wien Austria
OLML Oberösterreichisches Landesmuseum, Linz Austria
OUMNH University Museum of Natural History, Oxford United Kingdom
RMNH Nationaal Natuurhistorisch Museum Naturalis, Leiden The Netherlands
SEMC University of Kansas, Snow Entomological Museum, Lawrence United States
SMFD Forschungsinstitut und Naturmuseum Senckenberg, Frankfurt am Main Germany
SMNG Staatliches Museum für Naturkunde, Görlitz Germany
SMNK Staaliches Museum für Naturkunde, Karlsruhe Germany
SMNS Staatliches Museum für Naturkunde, Stuttgart Germany
SOFM National Museum of Natural History, Sofia Bulgaria
UASK Ukrainian Academy of Sciences, Institute of Zoology, Kiev Ukraine
USNM National Museum of Natural History, Washington United States
UWCP University of Wroclaw, Wroclaw Poland
UZIU Uppsala University, Uppsala Sweden
ZIN Russian Academy of Sciences, Zoological Institute, St. Petersburg Russia
ZMHB Museum für Naturkunde der Humboldt-Universität, Berlin Germany
ZMUC University of Copenhagen, Zoological Museum, Copenhagen Denmark
ZMUM Moscow State University, Moscow Russia
ZSM Zoologische Staatssammlung, München Germany
Geographic distribution
The Palaearctic faunal region comprises the main continental landmasses of temperate Eurasia, the Arabian Peninsula and northern Africa, as well as peripheral archipelagos such as the Azores in the west and Japan to the east. For pragmatic reasons, the precise delimitation of the Palaearctic region applied in this catalogue as well as the classification into the four rather artificial subregions Europe, Northern Africa, Northern Asia and Southwestern Asia (Figure 1) and the assessed geographical units listed below are mainly political. However, biogeographically isolated island groups such as the Canary Islands are recorded separately, even if they are not politically independent. As a shorthand, we apply the widely used international license plate codes for the areas representing independent countries. Following Lelej (2002), the large areas of Russia (RUS) and China (RC) were additionally subdivided into six and seven areas respectively (Figure 2, 3). Similarly, isolated islands or archipelagos were also attributed distinct codes as for instance E(Ca) for the Canary Islands, politically part of Spain (E). Spelling of the standardized geographical entities conforms to the Times Comprehensive Atlas of the World (11th edn, 2003). The current country of historical type collection localities was inferred by reference to the Times Atlas of World History (4th edn, 1993) in cases of doubt. We treat all indigenous taxa known to occur in this region. Most problematic in this respect are the transition zone of the Himalayan range and southern China. There are several taxa described from northwestern India not yet recorded from the Palaearctic region as adopted in this catalogue but which might be expected to occur there (Gupta, 1993).
The distributional data given for each species are based on the surveyed literature supplemented with considerable unpublished data derived from our osmiine bee database. Literature data were excluded if there was a well-founded suspicion that the species identification was incorrect. Distributional data that are assumed to be doubtful (i.e. deserving confirmation) are indicated with a question mark. Given the difficulties in identifying osmiine bees due to the poor taxonomic state and the lack of comprehensive keys, the current distributional data are expected to be incomplete and to contain errors.
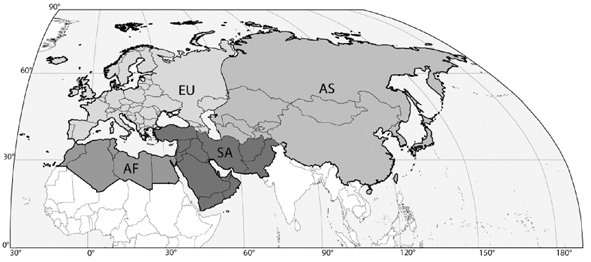
Figure 1.Map of the Palaearctic region and its four subregions as used in this catalogue. EU: Europe. AF: Northern Africa. AS: Northern Asia. SA: Southwestern Asia.
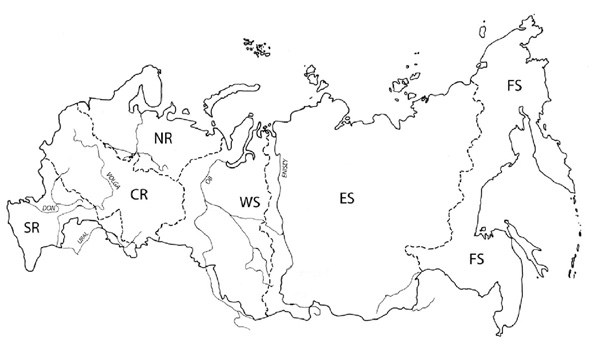
Figure 2. Map of Russia and its six subunits as used in this catalogue. SR: Southern European Russia. CR: Central European Russia. NR: Northern European Russia. WS: Western Siberia. ES: Eastern Siberia. FS: Far Eastern Siberia. Map adapted from Lelej (2002).
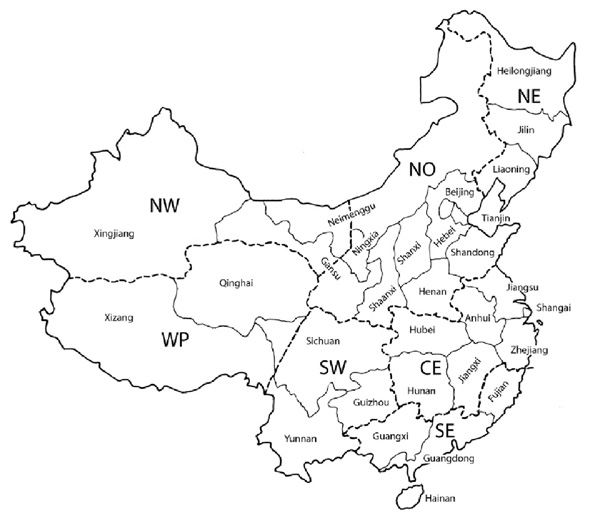
Figure 3. Map of China (excluding Taiwan) and its seven subunits as used in this catalogue. NW: Northwestern China. NO: Northern China. NE: Northeastern China. WP: Western Chinese Plateau. SW: Southwestern China. CE: Central China. SE: Southeastern China. Map adapted from Lelej (2002).
Geographic areas used in this catalogue
Biogeographic regions of the world:
Afr Afrotropic
Aus Australian
Nea Nearctic
Neo Neotropic
Ori Oriental
Pal Palaearctic
Subregions of the Palaearctic:
AF Northern Africa
AS Northern Asia
EU Europe
SA Southwestern Asia
Countries of the Palaearctic:
A (EU) Austria
AFG (SA) Afghanistan
AL (EU) Albania
AND (EU) Andorra
ARM (EU) Armenia
AZ (EU) Azerbaijan
B (EU) Belgium
BG (EU) Bulgaria
BIH (EU) Bosnia and Herzegovina
BRN (SA) Bahrain
BY (EU) Belarus
CH (EU) Switzerland
CY (SA) Cyprus
CZ (EU) Czech Republic
D (EU) Germany
DK (EU) Denmark
DK(Fa) (EU) Faroe Islands
DZ (AF) Algeria
E (EU)/(AF) Spain
E(Ba) (EU) Balearic Islands
E(Ca) (AF) Canary Islands
EST (EU) Estonia
ET (AF) Egypt
F (EU) France
F(Co) (EU) Corsica
FIN (EU) Finland
FL (EU) Liechtenstein
GB (EU) United Kingdom
GE (EU) Georgia
GR (EU) Greece
GR(Cr) (EU) Crete
H (EU) Hungary
HR (EU) Croatia
I (EU) Italy
I(Sa) (EU) Sardinia
I(Si) (EU) Sicily
IL (SA) Israel and Palestine
IR (SA) Iran
IRL (EU) Ireland
IRQ (SA) Iraq
IS (EU) Iceland
J (AS) Japan
JOR (SA) Jordan
KP (AS) North Korea
KS (AS) Kyrgyzstan
KWT (SA) Kuwait
KZ (EU)/(AS) Kazakhstan
L (EU) Luxembourg
LAR (AF) Libya
LT (EU) Lithuania
LV (EU) Latvia
M (EU) Malta
MA (AF) Morocco
MD (EU) Moldova
MGL (AS) Mongolia
MK (EU) Macedonia
N (EU) Norway
NL (EU) Netherlands
OM (SA) Oman
P (EU)/(AF) Portugal
P(Az) (EU) Azores
P(Ma) (AF) Madeira
PAK (SA) Pakistan
PL (EU) Poland
Q (SA) Qatar
RC (AS) China
RC(CE) (AS) Central China
RC(NE) (AS) Northeastern China
RC(NO) (AS) Northern China
RC(NW) (AS) Northwestern China
RC(SE) (AS) Southeastern China
RC(SW) (AS) Southwestern China
RC(WP) (AS) Western Chinese Plateau
RL (SA) Lebanon
RO (EU) Romania
ROK (AS) South Korea
RUS (EU)/(AS) Russia
RUS(CR) (EU) Central European Russia
RUS(ES) (AS) Eastern Siberia
RUS(FS) (AS) Far Eastern Siberia
RUS(NR) (EU) Northern European Russia
RUS(SR) (EU) Southern European Russia
RUS(WS)(AS) Western Siberia
S (EU) Sweden
SA (SA) Saudi Arabia
SCG (EU) Serbia and Montenegro
SK (EU) Slovakia
SLO (EU) Slovenia
SYR (SA) Syria
TJ (AS) Tajikistan
TM (AS) Turkmenistan
TN (AF) Tunisia
TR (EU)/(SA) Turkey
TW (AS) Taiwan
UA (EU) Ukraine
UAE (SA) United Arab Emirates
UZ (AS) Uzbekistan
Y (SA) Yemen
Y(So) (SA) Soqotra
Additionally, the following more loosely defined or border-crossing geographic terms covering certain mountain ranges, peninsulas and deserts are used if a current country could not be attributed with certainty to the locality information given in the literature:
Arabia (SA) for “Arabian Peninsula”, “Arabie”, “Arabien”
Balkans (EU)for “Balkanhalbinsel”, “Balkans”, “Dalmatia”, “Jugoslawien”, “Yugoslavia”
Caucasus (EU) for “Caucasus”, “Kaukasus”, “Transcaucasia”, “Transkaukasien”
Central Asian Ranges (AS) for “Hindu Kush”, “Karakoram”, “Pamir”, “Tien Shan”
Korea (AS) for “Corée”, “Korea”, “Korean Peninsula”
Sahara (AF)for “North African Desert”, “Sahara”
Turkestan (AS) for “Central Asia”, “Karakum Desert”, “Kyzylkum Desert”, “Mittelasien”, “Transcaspia”, “Turcestanica”, “Turkestan”
Species identification
To facilitate the identification of the Palaearctic osmiine bees, we cite for each species all literature sources in which it is described, compared with similar species or contained in keys. We do not cite the reference containing the original description under this section, though the original description often includes extensive descriptions or keys. Instead, these references are cited after each available species-group name.
Nesting behaviour
The description of the nesting behaviour is based on an extensive literature study on osmiine nesting biology as well as on own field observations (A. Müller, C. Praz, J. Neff, G. LeGoff and C. Sedivy, unpublished).
Flower preferences
The data on flower preferences are based on a compilation of flower records from the literature (A. Müller, unpublished) as well as on results of ongoing pollen-analytical studies at the Entomological Collection of the ETH Zürich.